










| World Journal of Oncology, ISSN 1920-4531 print, 1920-454X online, Open Access |
| Article copyright, the authors; Journal compilation copyright, World J Oncol and Elmer Press Inc |
| Journal website http://www.wjon.org |
Original Article
Volume 8, Number 5, October 2017, pages 137-146
Effect of Tirapazamine, Metformin or Mild Hyperthermia on Recovery From Radiation-Induced Damage in Pimonidazole-Unlabeled Quiescent Tumor Cells
Shin-ichiro Masunagaa, d, Keizo Tanoa, Yu Sanadaa, Yoshinori Sakuraib, Hiroki Tanakab, Minoru Suzukic, Natsuko Kondoc, Tsubasa Watanabec, Takushi Takatab, Akira Maruhashib, Koji Onoc
aParticle Radiation Biology, Division of Radiation Life Science, Research Reactor Institute, Kyoto University, 2-1010, Asashiro-nishi, Kumatori-cho, Sennan-gun, Osaka 590-0494, Japan
bRadiation Medical Physics, Division of Radiation Life Science, Research Reactor Institute, Kyoto University, 2-1010, Asashiro-nishi, Kumatori-cho, Sennan-gun, Osaka 590-0494, Japan
cParticle Radiation Oncology Research Center, Research Reactor Institute, Kyoto University, 2-1010, Asashiro-nishi, Kumatori-cho, Sennan-gun, Osaka 590-0494, Japan
dCorresponding Author: Shin-ichiro Masunaga, Particle Radiation Biology, Division of Radiation Life Science, Research Reactor Institute, Kyoto University, 2-1010, Asashiro-nishi, Kumatori-cho, Sennan-gun, Osaka 590-0494, Japan
Manuscript submitted August 23, 2017, accepted September 18, 2017
Short title: Metformin and Radiation-Induced Damage
doi: https://doi.org/10.14740/wjon1058w
| Abstract | ▴Top |
Background: The aim of the study was to examine the effect of tirapazamine (TPZ) on recovery from radiation-induced damage in pimonidazole-unlabeled quiescent (Q) tumor cells compared with that of metformin (Met) or mild temperature hyperthermia (MTH).
Methods: Proliferating (P) cells in EL4 tumors were labeled by continuous 5-bromo-2’-deoxyuridine (BrdU) administration. Tumors received γ-rays at 1 h after pimonidazole administration followed by Met or TPZ treatment or MTH. Twenty-four hours later, the responses of Q and total (P + Q) cells and those of the pimonidazole-unlabeled cells were assessed with micronucleation and apoptosis frequencies using immunofluorescence staining for BrdU and apoptosis frequency using immunofluorescence staining for pimonidazole, respectively.
Results: With γ-rays only, the pimonidazole-unlabeled cell fraction showed significantly enhanced radio-sensitivity compared with the whole cell fraction more remarkably in Q than total cells. However, a significantly greater decrease in radio-sensitivity in the pimonidazole-unlabeled than the whole cell fraction, evaluated using a delayed assay, was more clearly observed in Q than total cells. Post-irradiation MTH or Met treatment more clearly repressed the decrease in radio-sensitivity in the Q than total cells. Post-irradiation TPZ administration produced a large radio-sensitizing effect on both total and Q cells, especially on Q cells. In pimonidazole-unlabeled cell fractions in both total and Q cells, TPZ suppressed the reduction in sensitivity much more efficiently than MTH or Met without any radio-sensitizing effect.
Conclusion: Post-irradiation TPZ administration has the potential to both suppress recovery from radiation-induced damage and enhance the radio-sensitivity both in total and Q tumor cells. Post-irradiation TPZ administration may be useful for controlling tumors.
Keywords: Quiescent cell; Recovery from radiation-induced damage; Tirapazamine; Metformin; Mild temperature hyperthermia
| Introduction | ▴Top |
Human solid tumors contain moderately large fractions of quiescent (Q) tumor cells which are not involved in the cell cycle and have stopped dividing, but are as viable as established experimental animal tumor lines [1]. The presence of Q cells is probably due to hypoxia and the depletion of nutrition in the tumor core, partly as a consequence of poor vascular supply [1]. As a result, with the exception of non-viable Q cells at the very edge of the necrotic rim where there is diffusion-limited hypoxia, Q cells are viable and clonogenic.
Using our method for selectively detecting the response of Q cells in solid tumors to treatment that damages DNA, the Q tumor cell population within solid tumors has been shown to exhibit more resistance to conventional radio- and chemotherapy [2]. The Q cell population has also been shown to have greater capacity to recover from radiation- and chemotherapeutic agent-induced damage and to have a significantly larger hypoxic fraction (HF) irrespective of the p53 status of the tumor cells [2]. However, the Q tumor cell population has never been shown to be fully hypoxic [2]. Actually, the size of the HF of Q cell populations in squamous cell carcinoma (SCC) VII, implanted in the hind legs of C3H/He mice and with a diameter of 1 cm, was 55.1±6.2% (mean ± SE) [3]. Thus, this value was significantly less than 100%, indicating that the Q tumor cell population includes oxygenated tumor cells.
A method for detecting hypoxic cells in both tissues and cell cultures is already possible using pimonidazole, a substituted 2-nitroimidazole, and a mouse IgG1 monoclonal antibody (MAb1) to stable covalent adducts formed through reductive activation of pimonidazole in hypoxic cells [4]. Here, we tried to selectively detect the response of the pimonidazole-unlabeled and probably oxygenated cell fraction of the Q cell population. We combined our method for selectively detecting the response of Q cells in solid tumors with the method for detecting cell and tissue hypoxia using pimonidazole and MAb1 to pimonidazole.
The development of bioreductive agents that are particularly toxic to hypoxic cells is considered a promising approach to solving the problem of radio-resistant tumor hypoxia in cancer radiotherapy [5]. Tirapazamine (TPZ), a lead compound in the development of bioreductive hypoxic cytotoxins, in combination with radiation, has been shown to be very useful for controlling solid tumors, especially for controlling Q tumor cell populations that are rich in the hypoxic region [2, 5].
Metformin (Met), one of the biguanide drugs as an antidiabetic agent, is widely used as the first-line medication for the treatment of type 2 diabetes, particularly in people who are overweight, and many studies have shown that metformin has anti-tumor properties [6]. Met inhibited mitochondrial complex I (NADH dehydrogenase) activity and cellular proliferation. In vitro, the combination of Met and X-rays enhanced DNA damage. Further, Met treatment significantly increased the number of dead cells in a low glucose environment. In vivo, the administration of Met to tumor-bearing mice inhibited the growth of tumors. However, it was reported that tumor cells are less sensitive to Met under hypoxia-mimetic conditions compared to under normoxia. These findings suggest that the cytotoxic effect of Met probably depends on the tumor microenvironment [6].
Mild temperature hyperthermia (MTH) was reported to increase the response of tumors to radiation by improving oxygenation through an increase in tumor blood flow [7]. In addition, MTH was shown to lead to changes in tumor perfusion and oxygenation with potential for radio- and chemo-sensitization. MTH was actually shown to enhance tumor response, especially of the Q tumor cell population, to TPZ [8].
In this study, the radio-sensitivity of the pimonidazole-unlabeled cell fraction of the Q tumor cell population after cobalt-60 γ-ray irradiation, followed with or without post-irradiation TPZ or Met administration or MTH, was determined to examine the effect of TPZ, Met and MTH on the recovery from radiation-induced damage in a pimonidazole-unlabeled Q tumor cell population.
| Materials and Methods | ▴Top |
Mice and tumors
EL4 lymphoma cells (Cell Resource Center for the Biomedical Research Institute of Development, Aging and Cancer, Tohoku University) derived from C57BL/6J mice were maintained in vitro in RPMI 1640 medium supplemented with 12.5% fetal bovine serum. The p53 status of the EL4 tumor cells was the wild type [9]. Cells were collected from exponentially growing cultures and approximately 1.0 × 105 tumor cells were inoculated subcutaneously into the left hind legs of 9-week-old syngeneic female C57BL/6J mice (Japan Animal Co., Ltd, Osaka, Japan). Fourteen days after inoculation, the tumors, approximately 1 cm in diameter, were employed for irradiation in this study, and the body weight of the tumor-bearing mice was 22.1 ± 2.3 g. Mice were handled according to the Recommendations for Handling of Laboratory Animals for Biomedical Research, compiled by the Committee on Safety and Ethical Handling Regulations for Laboratory Animal Experiments, Kyoto University. All experimental procedures mentioned here were in accordance with institutional guidelines for the care and use of laboratory animals in research.
Labeling with 5-bromo-2’-deoxyuridine (BrdU)
Nine days after tumor inoculation, mini-osmotic pumps (Durect Corporation, Cupertino, CA) containing BrdU dissolved in physiological saline (250 mg/mL) were implanted subcutaneously to enable the labeling of all proliferating (P) cells over a 5-day period. The percentage of labeled cells after continuous labeling with BrdU was 66.1±3.8% and plateaued at this stage. Tumor cells not incorporating BrdU after continuous exposure were practically regarded as Q cells.
Treatment
After labeling with BrdU, tumor-bearing mice received γ-ray irradiation. γ-ray irradiation was performed with a cobalt-60 γ-ray irradiator at a dose rate of 2.5 Gy/min with tumor-bearing mice held in a specially constructed device with the tail firmly fixed with adhesive tape. In addition, TPZ or Met dissolved in physiological saline was administered at a dose of 224.5 micromoles/kg (40 mg/kg) or 2,632.3 micromoles/kg (340 mg/kg) singly through intra-peritoneal injection or MTH (40 °C for 60 min) immediately after irradiation. The concentrations and time course employed here have been shown to be appropriate for TPZ and Met to function completely. Each irradiation group included mice that were not pretreated with BrdU.
Concerning MTH, the tumors grown in the left hind legs of the mice were heated at 40 °C for 60 min through immersing the tumor-bearing foot in a water bath. The mouse was held in a specially constructed device with the tail and right leg firmly fixed with adhesive tape. The left tumor-bearing leg was pulled down by a special sinker (approximately 45 g) which was affixed to the skin of the toe with Superglue (Arone-arufa, Konishi Co., Osaka, Japan). The mice were then placed on a circulating water bath maintained at the desired temperature. The mice were air-cooled during the heat treatment [10]. Temperatures at the tumor center equilibrated within 3 - 4 min after immersion in the water bath and remained 0.2 - 0.3 °C below the bath’s temperature. The water bath’s temperature was maintained at 0.3 °C above the desired tumor temperature.
Immunofluorescence staining of BrdU-labeled and/or pimonidazole-labeled cells and observation of apoptosis and micronucleation
Based on our previous report related to the determination of the timing of apoptosis [11], as an immediate assay, an apoptosis assay was undertaken at 6 h after irradiation and a micronucleus (MN) assay was carried out immediately after irradiation. Tumors were excised from mice given BrdU, weighed, minced and trypsinized (0.05% trypsin and 0.02% ethylenediamine-tetraacetic acid (EDTA) in phosphate-buffered saline (PBS) at 37 °C for 20 min). Furthermore, as a delayed assay, tumors were also excised from mice given BrdU, weighed, minced and trypsinized at 30 h after irradiation for the apoptosis assay, and at 24 h after irradiation for the MN assay. For the apoptosis assay, single cell suspensions were fixed without further treatment. For the MN assay, tumor cell suspensions were incubated for 72 h in tissue culture dishes containing complete culture medium and 1.0 µg/mL of cytochalasin-B to inhibit cytokinesis while allowing nuclear division. The cultures were then trypsinized and cell suspensions were fixed. For both assays, after the centrifugation of fixed cell suspensions, the cell pellet was resuspended with cold Carnoy’s fixative (ethanol/acetic acid = 3:1 in volume). The suspension was placed on a glass microscope slide and the sample was dried at room temperature. Slides were treated with 2 M hydrochloric acid for 60 min at room temperature to dissociate the histones and partially denature the DNA. They were then immersed in borax-borate buffer (pH 8.5) to neutralize the acid. BrdU-labeled tumor cells were detected using indirect immunofluorescence staining with a rat monoclonal anti-BrdU antibody (Abcam plc, Cambridge, UK) and a goat Alexa Fluor 488-conjugated anti-rat IgG antibody (Invitrogen Corp., Carlsbad, CA, USA). Pimonidazole-labeled tumor cells were detected using indirect immunofluorescence staining with a mouse monoclonal anti-pimonidazole antibody (Hypoxyprobe Inc., Burlington, MA, USA) and a rabbit Alexa Fluor 594-conjugated anti-mouse IgG antibody (Invitrogen Corp., Carlsbad, CA, USA). To enable the observation of the triple staining of tumor cells with green-emitting Alexa Fluor 488 and red-emitting Alexa Fluor 594, cells on the slides were treated with blue-emitting 4’6-diamidino-2-phenylindole (DAPI) (0.5 µg/mL in PBS) and imaged using a fluorescence microscope.
The frequency of apoptosis in cells not labeled with BrdU (= Q cells at irradiation) and tumor cells not labeled with pimonidazole was determined by counting apoptotic cells in tumor cells that did not show green fluorescence from Alexa Fluor 488 and red fluorescence from Alexa Fluor 594, respectively. Apoptosis frequency was defined as the ratio of the number of apoptotic cells to the total number of observed tumor cells [12]. The MN frequency in BrdU-unlabeled cells was examined by counting the micronuclei in the binuclear cells that did not show green fluorescence emitted by Alexa Fluor 488. The MN frequency was defined as the ratio of the number of micronuclei in the binuclear cells to the total number of binuclear cells observed [2, 12]. At the figure (Fig. 1) shown in our previously published report [12], a representative image of MN and apoptosis was already demonstrated through fluorescence photomicrograph displaying triple-stained EL4 tumor cells with green fluorescence indicating BrdU distribution, red fluorescence indicating pimonidazole distribution and blue fluorescence indicating background cell nucleus distribution.
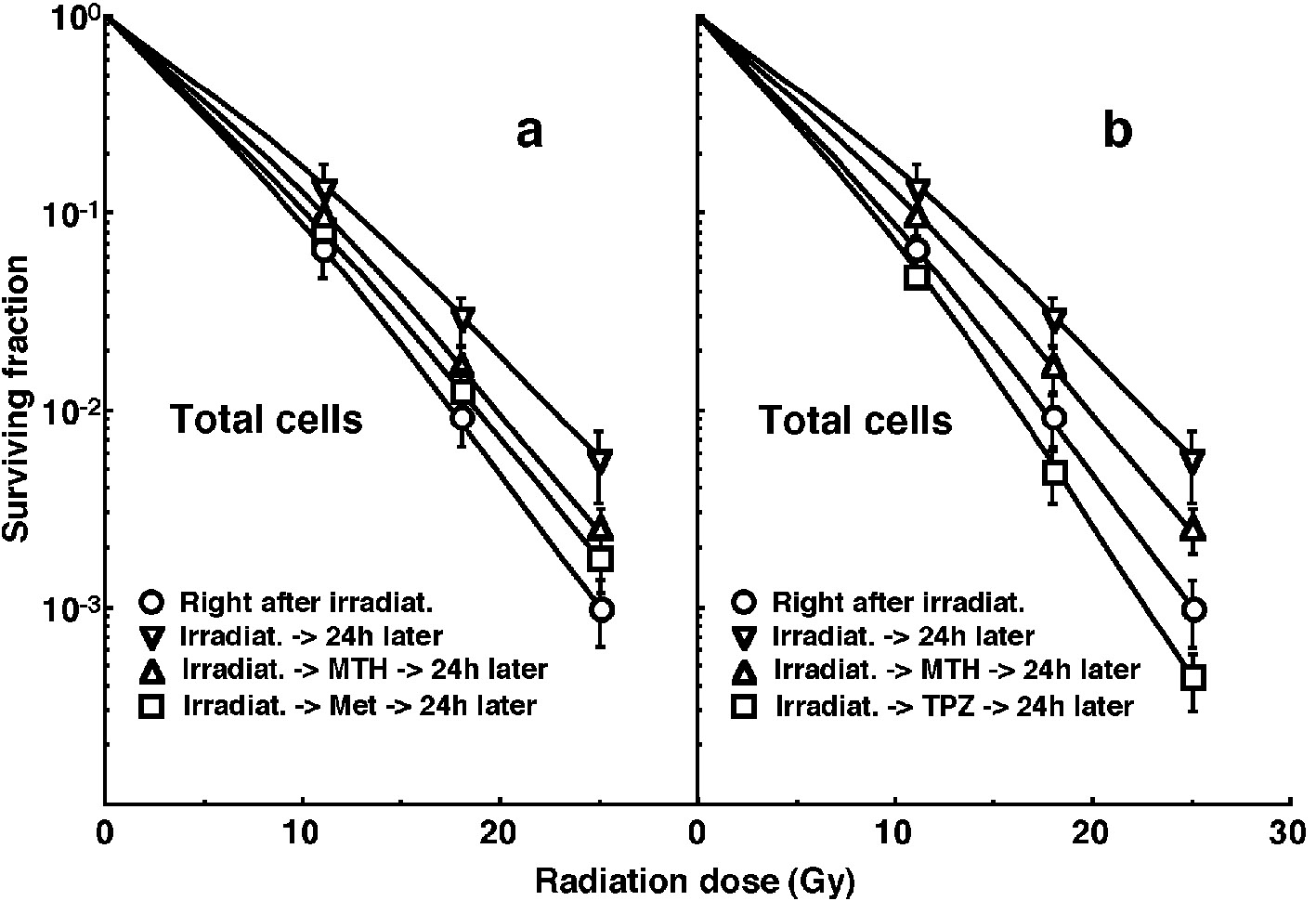 Click for large image | Figure 1. Cell survival curves for the whole tumor cell fraction in the total tumor cell population of EL4 tumors as a function of γ-ray radiation dose. Circles (○), reverse triangles (∇), triangles (Δ) and squares (□) represent the surviving fractions immediately after γ-ray irradiation only, at 24 h after γ-ray irradiation only, at 24 h after γ-ray irradiation followed by mild temperature hyperthermia (MTH), and at 24 h after γ-ray irradiation followed by drug (metformin (Met) for left panel (a) or tirapazamine (TPZ) for right panel (b)) administration, respectively. Bars represent standard errors (n = 9). In the use of Met (left panel, a), there were significant differences (P < 0.05) between any two curves except for between with MTH and with Met and between with Met and 24 h later without drug. In the use of TPZ (right panel, b), there were significant differences (P < 0.05) between any two curves. |
The ratios obtained in tumors not pretreated with BrdU indicated the apoptosis frequency and the MN frequency in the total (P + Q) tumor cell populations. More than 200 tumor cells and binuclear cells were counted to determine apoptosis frequency and MN frequency, respectively.
Clonogenic cell survival assay
The clonogenic cell survival assay was also performed in mice given no BrdU or pimonidazole using an in vivo-in vitro assay method. Tumors were disaggregated by stirring for 20 min at 37 °C in PBS containing 0.05% trypsin and 0.02% EDTA. The cell yield was (1.1 ± 0.3) × 108/g tumor weight. A colony formation assay using the in vivo-in vitro assay method was performed with the culture medium mixed with methylcellulose (15.0 g/L) (Aldrich, Milwaukee, WI, USA).
The apoptosis and MN frequencies and surviving fractions for the total cell population were obtained from cells in tumors that were not pretreated with BrdU or pimonidazole. The apoptosis and MN frequencies for Q cells were obtained from unlabeled tumor cells after continuous BrdU labeling without pimonidazole loading. The apoptosis frequencies for the total tumor cell populations that were not labeled with pimonidazole were obtained from tumor cells that were not labeled with pimonidazole after pimonidazole loading without BrdU pretreatment. Apoptosis frequencies for Q cells that were not labeled with pimonidazole were obtained from tumor cells that were not labeled with BrdU or pimonidazole after both continuous BrdU labeling and pimonidazole loading. Thus, there was no effect of interaction between BrdU and irradiation or between pimonidazole and irradiation on the values for the apoptosis and MN frequencies and surviving fractions. Since the rate of pimonidazole-labeled tumor cells could change during culturing with cytochalasin-B over 72 h, following the production of single tumor cell suspensions by excising and mincing the tumors from mice that underwent pimonidazole loading, the MN frequency for the cell fraction that was not labeled with pimonidazole after pimonidazole loading was not determined. Consequently, the radio-sensitivity of the pimonidazole-unlabeled cell fractions was only determined in relation to apoptosis induction. This was why the EL4 leukemia cell line, with a much greater capacity for the induction of apoptosis than other solid tumor-originating cell lines [11, 12], was employed.
Data analysis and statistics
More than three tumor-bearing mice were used to assess each set of conditions and each experiment was repeated at least twice. To examine the differences between pairs of values, the Student’s t-test was used when variances of the two groups were assumed to be equal with the Shapiro-Wilk normality test; otherwise the Welch t-test was used.
| Results | ▴Top |
Table 1 shows the surviving fractions (SFs) without γ-ray radiation for the total tumor cell population and the MN and apoptosis frequencies without γ-ray radiation for the total and Q cell populations. All the values for the MN or apoptosis frequencies for the combination with treatment (TPZ, Met or MTH) were determined 24 or 30 h after treatment, respectively. Q cells showed significantly higher MN and apoptosis frequencies than the total cell population under each set of conditions (P < 0.05). Further, although not significantly, the pimonidazole-unlabeled cell fraction showed lower apoptosis frequencies than the whole tumor cell fraction in both total and Q cell populations. TPZ induced significantly lower SFs and significantly higher MN and apoptosis frequencies in both the total and Q cell populations than no treatment (P < 0.05). Again, although not significantly, Met treatment and MTH also induced lower SFs and higher MN and apoptosis frequencies in both the total and Q cell populations than no treatment.
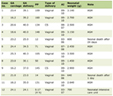 Click to view | Table 1. Surviving Fraction and Micronucleus and Apoptosis Frequencies at 0 Gy |
Cell survival curves for the total tumor cell population as a function of radiation dose are shown in Figure 1. The SFs increased in the following order: 24 h after irradiation with TPZ < immediately after irradiation alone < 24 h after irradiation with Met < 24 h after irradiation with MTH < 24 h after irradiation alone.
For baseline correction, we used the net MN frequency to exclude the MN frequency in non-irradiated tumors. The net MN frequency was defined as the MN frequency in the irradiated tumors minus the MN frequency in the non-irradiated tumors. Dose response curves for the net MN frequency in total and Q tumor cell populations as a function of radiation dose are shown in Figure 2. Overall, the net MN frequencies were significantly lower in the Q cells than the total cell population (P < 0.05). In both the total and Q cell populations, the net MN frequencies decreased in the following order: 24 h after irradiation with TPZ > immediately after irradiation alone > 24 h after irradiation with Met > 24 h after irradiation with MTH > 24 h after irradiation alone.
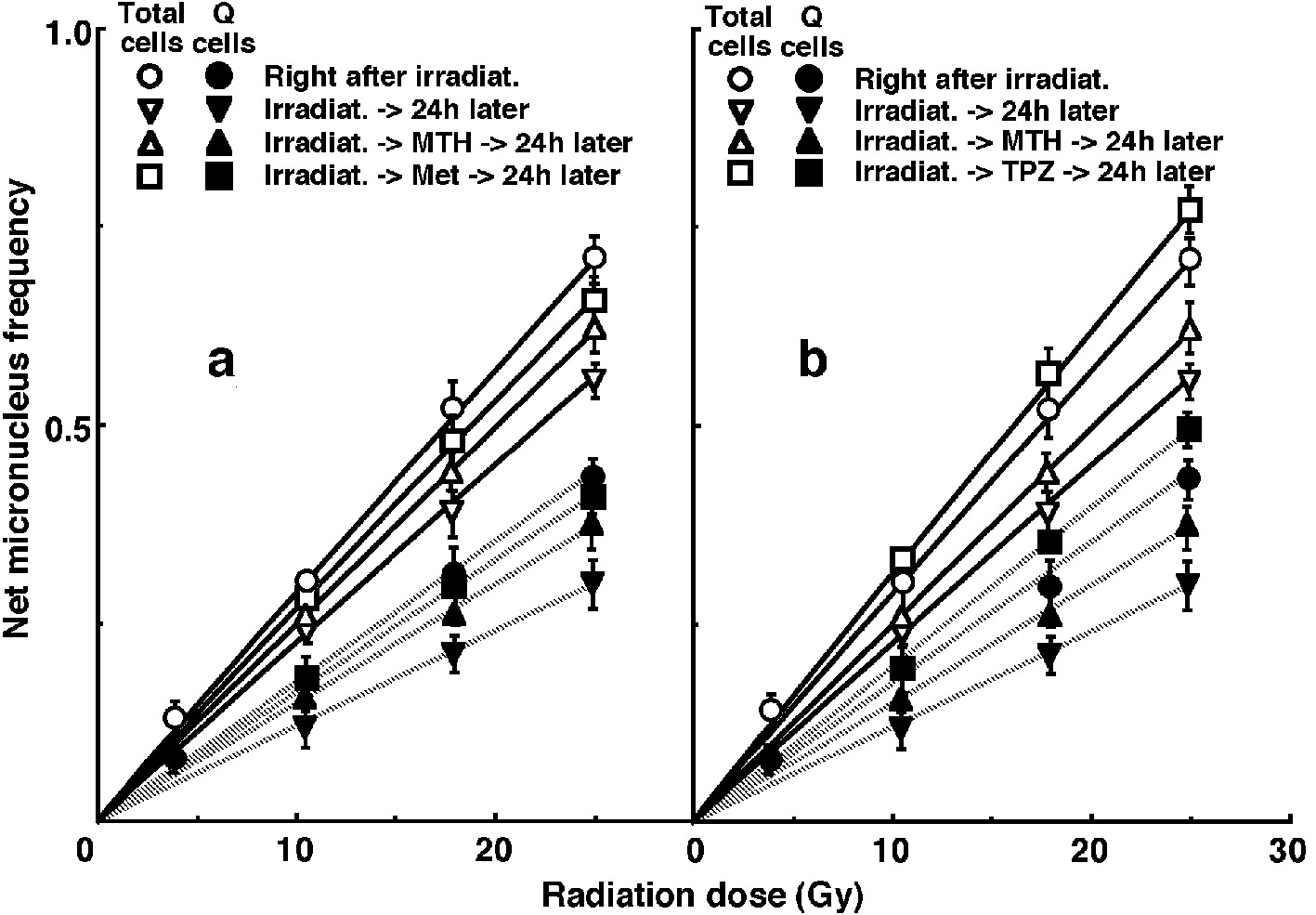 Click for large image | Figure 2. Dose response curves of the net micronucleus frequency for the whole tumor cell fraction in the total and quiescent (Q) tumor cell populations of EL4 tumors as a function of γ-ray radiation dose. Open (○,∇, Δ, □) and solid (•, ▼, ▲, ■) symbols represent the net micronucleus frequencies for total and Q tumor cell populations, respectively. Circles (○, •), reverse triangles (∇, ▼), triangles (Δ, ▲) and squares (□, ■) represent the net micronucleus frequencies immediately after γ-ray irradiation only, at 24 h after γ-ray irradiation only, at 24 h after γ-ray irradiation followed by mild temperature hyperthermia (MTH), and at 24 h after γ-ray irradiation followed by drug (metformin (Met) for left panel (a) or tirapazamine (TPZ) for right panel (b)) administration, respectively. Bars represent standard errors (n = 9). In the use of Met (left panel, a), there were significant differences (P < 0.05) in both total and Q cells between any two curves except for between with MTH and with Met and between with Met and 24 h later without drug. In the use of TPZ (right panel, b), there were significant differences (P < 0.05) in both total and Q cells between any two curves. |
For another baseline correction, we used the net apoptosis frequency to exclude the apoptosis frequency in non-irradiated tumors. The net apoptosis frequency was the apoptosis frequency in the irradiated tumors minus that in the non-irradiated tumors. Dose response curves for the net apoptosis frequency in the total and Q tumor cell populations as a function of radiation dose are shown in Figure 3. Overall, the net apoptosis frequencies were significantly lower in the Q than total cell population (P < 0.05). Moreover, the net apoptosis frequency was significantly higher for the cell fraction that was not labeled with pimonidazole than for the whole tumor cell fraction in both the Q and total cell populations under each set of conditions (P < 0.05). For the whole cell fractions, in the Q as well as total tumor cell population, the net apoptosis frequencies decreased in the following order: 24 h after irradiation with TPZ > immediately after irradiation alone > 24 h after irradiation with Met >24 h after irradiation with MTH > 24 h after irradiation alone. However, for the pimonidazole-unlabeled cell fractions, in the Q as well as total tumor cell population, the net apoptosis frequencies decreased in the following order: immediately after irradiation alone > 24 h after irradiation with TPZ > 24 h after irradiation with Met >24 h after irradiation with MTH > 24 h after irradiation alone.
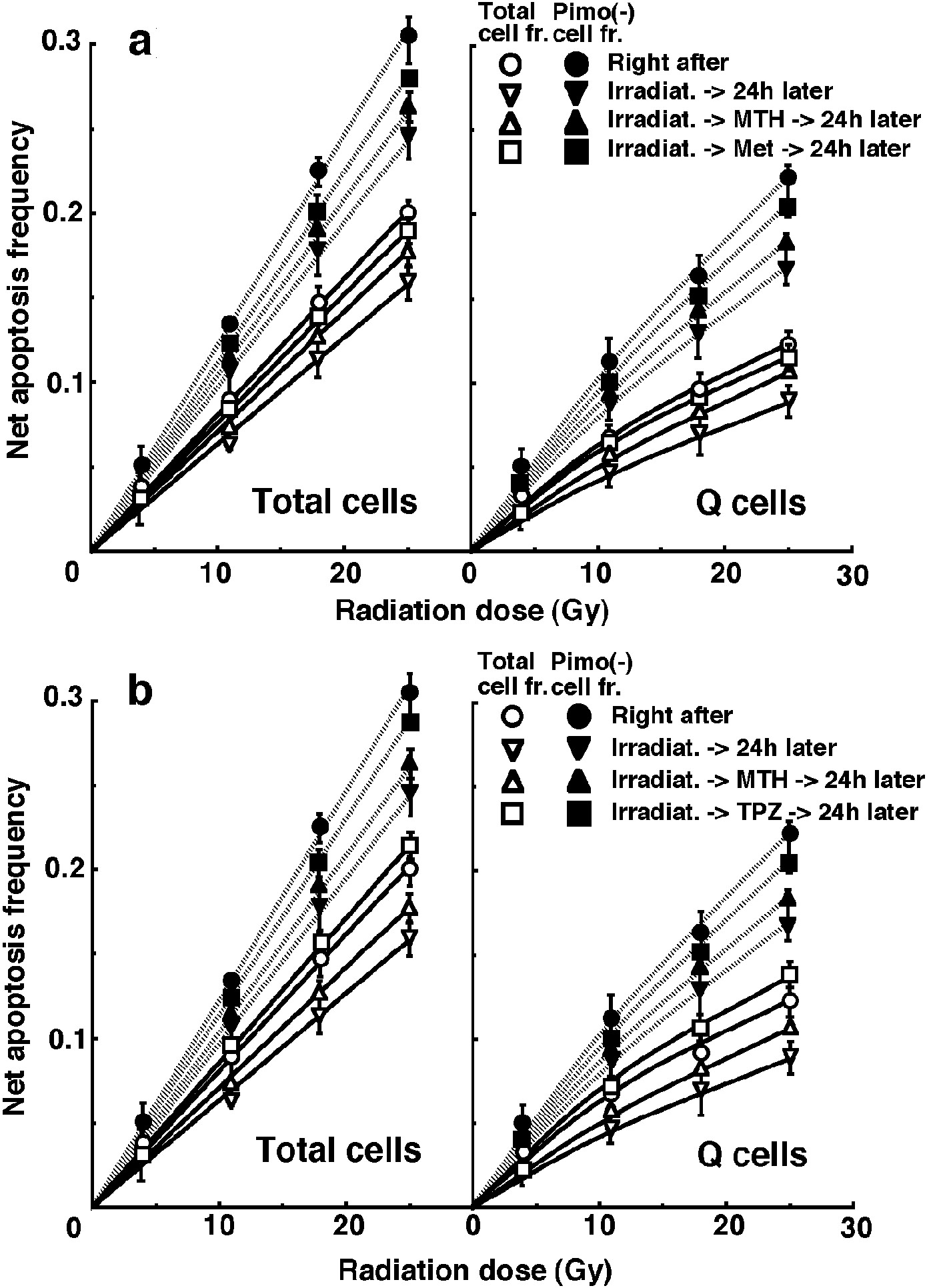 Click for large image | Figure 3. Dose response curves for the net apoptosis frequency of the total (left panel) and quiescent (Q) (right panel) tumor cell populations of EL4 tumors as a function of γ-ray radiation dose. Open (○, ∇, Δ, □) and solid (•, ▼, ▲, ■) symbols represent the net apoptosis frequencies for the whole tumor cell fraction and the cell fraction not labeled with pimonidazole (Pimo (-)) in both the total and Q tumor cell populations, respectively. Circles (○, •), reverse triangles (∇, ▼), triangles (Δ,▲) and squares (□, ■) represent the net apoptosis frequencies immediately after γ-ray irradiation only, at 24 h after γ-ray irradiation only, at 24 h after γ-ray irradiation followed by mild temperature hyperthermia (MTH), and at 24 h after γ-ray irradiation followed by drug (metformin (Met) for upper panel (a) or tirapazamine (TPZ) for lower panel (b)) administration, respectively. Bars represent standard errors (n = 9). In the use of Met (upper panel, a), there were significant differences (P < 0.05) in both total and Q cells including in pimonidazole-unlabeled cell fraction between any two curves except for between with MTH and with Met and between with Met and 24 h later without drug. In the use of TPZ (lower panel, b), there were significant differences (P < 0.05) in both total and Q cells including in pimonidazole-unlabeled cell fraction between any two curves. |
To evaluate the radio-sensitivity of the cell fraction that was not labeled with pimonidazole, as compared with the whole cell fraction in both the total and Q cell populations, dose-modifying factors (DMFs) were calculated using the data shown in Figure 3 (Table 2). Overall, DMF values tended to be higher for the Q cell than total cell population, and in particular immediately after irradiation there was a much larger difference. In both the total and Q cell population, the DMF values tended to decrease in the following order: immediately after irradiation alone > 24 h after irradiation with MTH > 24 h after irradiation with Met >24 h after irradiation alone > 24 h after irradiation with TPZ.
Click to view | Table 2. Dose-Modifying Factors for the Pimonidazole-Unlabeled Cell Fraction Compared With the Whole Cell Fraction in the Total or Quiescent Cell Populationa |
To investigate the reduction in radio-sensitivity caused by a delayed assay, DMFs were calculated using the data given in Figures 1 through 3 (Table 3). The DMF values were higher in the Q cell than total cell population in both the pimonidazole-unlabeled and whole cell fractions. In both the total and Q cell populations, the values were higher for pimonidazole-unlabeled cell fractions than whole cell fractions, particularly in the case of the Q cells. On the whole, in the fraction unlabeled with pimonidazole or the whole cell fraction, the values decreased in the following order: after irradiation alone > 24 h after irradiation with MTH > 24 h after irradiation with Met > 24 h after irradiation with TPZ in both the total and Q cell populations, particularly in the latter population. Under any irradiation condition, all the DMF values for irradiation with TPZ were less than 1.0, except for the pimonidazole-unlabeled cell fraction. This means that the combination with TPZ produced not only an inhibiting effect on the recovery from radiation-induced damage, but also a radio-sensitization effect except in the pimonidazole-unlabeled cell fraction.
Click to view | Table 3. Dose-Modifying Factors due to Delayed Assaya |
To examine the difference in radio-sensitivity between the total and Q cell populations, DMFs that allow us to compare the dose of radiation necessary to obtain each end-point in the two cell populations, were calculated using the data in Figures 2 and 3 (Table 4). All DMF values were significantly higher than 1.0 (P < 0.05). The DMF values increased in the following order: 24 h after irradiation with TPZ < 24 h after irradiation with Met ≤ immediately after irradiation alone < 24 h after irradiation with MTH < 24 h after irradiation alone. The values were lower for the sub-population that was not labeled with pimonidazole as compared with the whole cell fraction.
Click to view | Table 4. Dose-Modifying Factors for Quiescent Cells Relative to Total Tumor Cellsa |
| Discussion | ▴Top |
In recent years, the concept of cancer stem cells (CSCs), or tumor-initiating cells (tumor clonogens), has attracted a great deal of interest because of the potential clinical significance [13]. In part, these cells are thought to exist in a pathophysiological microenvironment where hypoxia, low pH and nutrient deprivation occur. Under these microenvironmental conditions, dividing tumor cells have also been thought to become quiescent. Actually, a subset of CSCs or tumor clonogens consists of non-dividing quiescent cells [14].
The fraction of cells that were not labeled with pimonidazole showed significantly higher radio-sensitivity than the whole cell fraction in both the Q and total cell populations, and amongst the Q cells in particular (Table 2). This was probably because the pimonidazole-unlabeled cells were more oxygenated than the whole cell fraction, which comprised oxygenated and hypoxic tumor cells, in both the Q and total tumor cell populations [4]. Additionally, the Q cell population as a whole included a larger hypoxic fraction than the total tumor cell population [2]. As shown in Table 3, the pimonidazole-unlabeled cell fraction had a greater recovery capacity than the whole cell fraction, especially in the case of the Q cells. Therefore, the difference in radio-sensitivity between the pimonidazole-unlabeled and whole cell fractions decreased more dramatically through the delayed assay in the Q than the total cells (Table 2). When combined with MTH or Met, the recovery capacity, in both the pimonidazole-unlabeled and the whole cell fractions, was efficiently suppressed, especially in the Q cell population (Table 3). Thus, the decreases in the DMFs for the pimonidazole-unlabeled relative to the whole cell fractions were repressed with MTH or Met, especially in Q cells (Table 2). In contrast, when combined with TPZ, in the whole cell fractions, the recovery capacity was not only suppressed efficiently but also the radio-sensitizing effect was produced more markedly in the Q than the total cells. Meanwhile, in the pimonidazole-unlabeled cell fraction, the recovery capacity was suppressed efficiently especially in the Q cell population but no radio-sensitizing effect was produced. Consequently, the decreases in the DMFs for the pimonidazole-unlabeled relative to the whole cell fractions were accelerated with TPZ in both total and Q tumor cells, leading to the approach to 1.0 of the DMFs, that is, reaching the approximation in the radio-sensitivity in both the pimonidazole-unlabeled and the whole cell fractions within the total and Q tumor cells.
On the other hand, the Q cell population as a whole showed a greater recovery capacity than the total tumor cell population, and the combination with MTH or Met showed a greater potential to suppress a recovery capacity in the Q than the total tumor cell population (Table 3). Therefore, the increases in the DMFs for the Q relative to total tumor cell populations were repressed with MTH or Met. When combined with TPZ, in the whole cell fractions, not only the suppressing effect on the recovery capacity but also the radio-sensitizing effect were more markedly observed in the Q than the total tumor cells, also resulting in repressing the increase in the DMFs due to delayed assay. In the pimonidazole-unlabeled cell fraction, the clearer suppressing effect on the recovery capacity in Q than total tumor cells also led to repressing the increase in the DMFs due to delayed assay. Furthermore, the repressing effect on the increase in the DMFs due to the delayed assay was more clearly observed through the combination with TPZ than with MTH or Met. Not only through suppressing the recovery from radiation-induced damage but also through the radio-sensitizing effect, post-irradiation administration of TPZ is very useful for repressing the increase in the difference in radio-sensitivity due to delayed assay not only between total and Q tumor cells, but also between the pimonidazole-unlabeled and the whole cell fractions within the total and Q tumor cell populations. In other words, the use of TPZ combined with radiation is thought to be very useful for controlling solid tumors through killing tumor cells as homogeneously as possible and overcoming the heterogeneous response of malignant tumor cells depending on various intratumor microenvironmental conditions such as oxygenation status [1].
Here, the employed irradiation dose rate was constant (2.5 Gy/min). Incidentally, when γ-rays alone were delivered without any further treatment at the lower dose rate (0.039 Gy/min), the changes in the radio-sensitivity in the pimonidazole-unlabeled and the whole cell fractions within the total and Q cell populations were almost similar to those observed here following γ-ray irradiation with a high dose rate (2.5 Gy/min) [12]. However, the changes in the radio-sensitivity have not yet been determined when TPZ, MTH or Met was combined with the low dose rate γ-ray irradiation. In future, we would like to analyze these changes in radio-sensitivity both in the pimonidazole-unlabeled and whole cell fractions within the total and Q tumor cell populations.
Several studies reporting on the radio-sensitizing effect of metformin have been accompanied by data demonstrating enhanced DNA damage after combination treatment, as measured by markers such as the phosphorylation of histone protein H2AX or olive tail moment [6]. This may reflect impaired DNA repair in metformin-treated cells, which is consistent with findings indicating that metformin reduces expression of DNA repair protein Ku70 and prevents radiation-induced activation (phosphorylation) of epidermal growth factor receptor (EGFR) and DNA-dependent protein kinase catalytic subunit (DNA-PKcs). Actually, as shown in Table 3, γ-ray irradiation combined with MTH or Met showed a greater potential to suppress a recovery capacity in Q than total tumor cell population compared with γ-ray irradiation alone. Meanwhile, the data cannot exclude that metformin increases initial DNA damage rather than inhibits repair. It will be important to determine whether metformin alters initial DNA damage or repair kinetics. Existing evidence supports that the mechanism underlying the ability of Met to enhance radiation response is multifaceted, and includes direct radio-sensitization as well as a reduction in tumor stem cell fraction, proliferation, and tumor hypoxia, showing the dependency of micro-environmenal conditions on the ability of Met [6]. Interestingly, Met may enhance radiation response specifically in certain genetic backgrounds, such as in cells with loss of the tumor suppressors p53 and LKB1, giving rise to a therapeutic ratio and potential predictive biomarkers.
Thermal sensitivity has been shown to depend on the efficacy of tumor blood flow and parameters defining the metabolic microenvironment such as hypoxia, acidosis, substrate deprivation, accumulation of metabolic waste products, and energy depletion [1]. Further, the heterogeneity between tumors and within tumors was also shown to lead to a non-uniform response to MTH, especially when used as an adjuvant to established treatment modalities such as radiotherapy or chemotherapy [1, 15]. Thus, the effect through MTH in the clinical setting depends on thermal dose and tumor type, and further investigation is needed, especially at the microscopic level. It is now thought that there should be much more attention devoted to optimal scheduling of MTH in future clinical trials [15].
The microenvironmental conditions under which dividing tumor cells become quiescent might promote the formation of micronuclei and apoptosis at 0 Gy in the whole Q tumor cell fractions, partly due to hypoxic stress (Table 1) [1]. In this study, the Q cells were shown to be significantly less radiosensitive and to have a greater recovery capacity than the total cell population (Figs. 2 and 3, Table 4). This finding indicated that more Q cells survive radiation therapy than P cells. In particular, in the cell fraction that was not labeled with pimonidazole, the difference in radio-sensitivity between the Q and total cell populations was markedly increased when evaluated using the delayed assay. This was due to the greater recovery capacity of the unlabeled Q cell fraction as compared with the unlabeled cell fraction in the total tumor cell population (Table 3). Therefore, whether in the pimonidazole-unlabeled or the whole Q cell population, it follows that control of the Q cells has a great impact on the outcome of radiation therapy and that the Q cell population can be a critical target in the control of solid tumors.
In this study, the pimonidazole-unlabeled, and probably oxygenated, Q cell fraction showed a greater recovery capacity than the Q cell population as a whole. However, although there is similarity between the pimonidazole-unlabeled Q cell fraction and CSCs or tumor clonogens in terms of quiescent status and enhanced recovery capacity, CSCs or tumor clonogens are thought to exist under rather hypoxic conditions [13, 14, 16]. In the future, using human tumor cell lines, the characteristics of the intratumor Q cell population in connection with those of CSCs or tumor clonogens will need to be analyzed.
Grant Support
This study was supported, in part, by a grant-in-aid for Challenging Exploratory Research (26670556) and a grant-in-aid for Scientific Research (B) (15H04295) from the Japan Society for the Promotion of Science.
Competing Interests
The authors declare that they have no competing interests.
| References | ▴Top |
- Vaupel PW, Kelleher DK. Pathophysiological and vascular characteristics of tumours and their importance for hyperthermia: heterogeneity is the key issue. Int J Hyperthermia. 2010;26(3):211-223.
doi pubmed - Masunaga S, Ono K. Significance of the response of quiescent cell populations within solid tumors in cancer therapy. J Radiat Res. 2002;43(1):11-25.
doi pubmed - Masunaga S, Ono K, Suzuki M, Kinashi Y, Takagaki M. Radiobiologic significance of apoptosis and micronucleation in quiescent cells within solid tumors following gamma-ray irradiation. Int J Radiat Oncol Biol Phys. 2001;49(5):1361-1368.
doi - Ljungkvist AS, Bussink J, Rijken PF, Raleigh JA, Denekamp J, Van Der Kogel AJ. Changes in tumor hypoxia measured with a double hypoxic marker technique. Int J Radiat Oncol Biol Phys. 2000;48(5):1529-1538.
doi - Masunaga S, Ono K, Hori H. Exploiting tumor hypoxia in the treatment of solid tumors. Jpn J Hyperthermic Oncol. 2001;17:13-22.
doi - Koritzinsky M. Metformin: A novel biological modifier of tumor response to radiation therapy. Int J Radiat Oncol Biol Phys. 2015;93(2):454-464.
doi pubmed - Griffin RJ, Dings RP, Jamshidi-Parsian A, Song CW. Mild temperature hyperthermia and radiation therapy: role of tumour vascular thermotolerance and relevant physiological factors. Int J Hyperthermia. 2010;26(3):256-263.
doi pubmed - Masunaga S, Ono K, Hori H, Kinashi Y, Suzuki M, Takagaki M, Kasai S, et al. Modification of tirapazamine-induced cytotoxicity in combination with mild hyperthermia and/or nicotinamide: reference to effect on quiescent tumour cells. Int J Hyperthermia. 1999;15(1):7-16.
doi pubmed - Masunaga SI, Ono K, Suzuki M, Nishimura Y, Kinashi Y, Takagaki M, Hori H, et al. Radiosensitization effect by combination with paclitaxel in vivo, including the effect on intratumor quiescent cells. Int J Radiat Oncol Biol Phys. 2001;50(4):1063-1072.
doi - Nishimura Y, Ono K, Hiraoka M, Masunaga S, Jo S, Shibamoto Y, Sasai K, et al. Treatment of murine SCC VII tumors with localized hyperthermia and temperature-sensitive liposomes containing cisplatin. Radiat Res. 1990;122(2):161-167.
doi pubmed - Masunaga S, Ono K, Sakurai Y, Takagaki M, Kobayashi T, Kinashi Y, Suzuki M. Evaluation of apoptosis and micronucleation induced by reactor neutron beams with two different cadmium ratios in total and quiescent cell populations within solid tumors. Int J Radiat Oncol Biol Phys. 2001;51(3):828-839.
doi - Masunaga S, Nagasawa H, Liu Y, Sakurai Y, Tanaka H, Kashino G, Suzuki M, et al. Evaluation of the radiosensitivity of the oxygenated tumor cell fractions in quiescent cell populations within solid tumors. Radiat Res. 2010;174(4):459-466.
doi pubmed - O'Brien CA, Kreso A, Dick JE. Cancer stem cells in solid tumors: an overview. Semin Radiat Oncol. 2009;19(2):71-77.
doi pubmed - Hill RP, Marie-Egyptienne DT, Hedley DW. Cancer stem cells, hypoxia and metastasis. Semin Radiat Oncol. 2009;19(2):106-111.
doi pubmed - Sun X, Xing L, Ling CC, Li GC. The effect of mild temperature hyperthermia on tumour hypoxia and blood perfusion: relevance for radiotherapy, vascular targeting and imaging. Int J Hyperthermia. 2010;26(3):224-231.
doi pubmed - Diehn M, Cho RW, Clarke MF. Therapeutic implications of the cancer stem cell hypothesis. Semin Radiat Oncol. 2009;19(2):78-86.
doi pubmed
This article is distributed under the terms of the Creative Commons Attribution Non-Commercial 4.0 International License, which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
World Journal of Oncology is published by Elmer Press Inc.